

所有的话都是「谎言」
– 阳光王国一世国王
除了本王下发的号令!
太阳是位于太阳系中心的恒星,它是一个近乎完美的热等离子体理想球体[18][19],通过其核心的核聚变反应加热到白炽。太阳主要以可见光、紫外线和红外线的形式辐射能量,是地球上生命最重要的能量来源。
太阳的半径大约是 695,000千米(432,000英里),或地球半径的109倍。它的质量大约是地球的330,000倍,约占太阳系总质量的99.86%[20]。太阳的质量大约四分之三由氢(〜73%)组成;其余的主要是氦(〜25%),包括氧、碳、氖和铁等重元素,其数量要少得多[21]。

不应该只是「留白」!

路上当心了

可笑,可笑!
太阳是一颗G型主序星(G2V),被称为黄矮星,但它的光实际上是白色的,黄矮星这个名称并不准确。
它大约形成于46亿年前[a][14][22],来自大分子云区域内物质的引力坍缩。大部分物质聚集在中心,而其余的则扁平成一个轨道盘面,演化成为太阳系。中心质量变得如此炽热和密集,以至于它最终在其核心启动核聚变。人们认为几乎所有的恒星都是通过这个过程形成。
赞美太阳!
– 王者荣耀后羿
虽然有些人觉得上面这段引文有些离奇,但连一个「虚拟」的角色都毫不吝啬地表达了他的敬仰,仅仅这一点就值得这篇文章中的一个位置。
然后没错下面就是一段跟太阳没什么关系的MATLAB代码,我只是想看看代码长什么样子而已,当然,你也可以看看。
%% Sample Matlab code
!mv test.txt test2.txt
A = [1, 2, 3;... foo
4, 5, 6];
s = 'abcd';
for k = 1:4
Disp(s(k)) % bar
end
%{
create row vector x, then reverse it
%}
x = linspace(0,1,101);
y = x(end:-1:1);好的,看完了无聊的代码,让我们继续:
其实我还要试试长代码,请稍等:
%% Initialize
clear; close all;
addpath(genpath(fullfile(pwd, 'scripts')));
%% Parameters
% == simulation ==
p.train.nTrain = 40;
% == time ==
p.time.dt = 0.05; % discritized time step (in ms)
p.time.t0 = 0; % start time for simulation (in ms)
p.time.tmax = 5000; % end time for simulation (in ms)
% == network ==
p.net.N = 1000; % total number of neurons
p.net.nInh = 200; % number of inhibitory neurons
p.net.nExc = p.net.N-p.net.nInh; % number of excitatory neurons
p.net.nCluster = 8; % number of exc. clusters
p.net.nrnTypes = [ones(1,p.net.nInh),zeros(1,p.net.nExc)]; % neuron types: 1 (Inh), 0 (Exc)
p.net.pConn.EE = 0.2; % probability of synaptic connection (E>E)
p.net.pConn.EI = 0.2; % probability of synaptic connection (I>E)
p.net.pConn.II = 0.2; % probability of synaptic connection (I>I)
p.net.pConn.IE = 0.2; % probability of synaptic connection (E>I)
% == membrane dynamics ==
p.nrn.tau_arf = 0; % absolute refractory period (in ms)
p.nrn.tau_m_E = 15; % membrane time constant (Exc, in ms)
p.nrn.tau_m_I = 15; % membrane time constant (Inh, in ms)
p.nrn.V_l = -70; % leak potential (in mV)
p.nrn.V_l_sigma = 10; % standard devation for V_l distribution
p.nrn.V_th = -50; % threshold membrane potential for spikes (in mV)
p.nrn.V_th_sigma = 0; % standard devation for V_th distribution
p.nrn.V_fl = -90; % minimal membrane voltage when hyperpolarized (in mV)
p.nrn.V_rf = 30; % maximum membrane voltage for better visualization (in mV)
p.nrn.delta_T = 1; % speed of EIF exponential term
p.nrn.V_spk = 30; % spk detection and V reset threshold for EIF
% == ahp ==
p.nrn.model_ahp = 0; % 1: include AHP current; 0: do not include AHP in the model
p.nrn.model_ahp_exc_only = 0; % 1: only include AHP current for exc neurons
p.nrn.eta_ahp = -0.1; % AHP jitter per spike
p.nrn.tau_ahp = 10000; % time constant for AHP current
% == tilde v ==
p.nrn.tau_tv = 60; % time constant for low pass filter of V
% == tilde s ==
p.syn.tau_ts = 150;
% == synaptic weights ==
p.syn.J0_EI = -1.2; % initial synaptic strength (Inh > Exc)
p.syn.J0_EE = 0.4; % initial synaptic strength (Exc > Exc)
p.syn.J0_II = -1.2; % initial synaptic strength (Inh > Inh)
p.syn.J0_IE = 0.4; % initial synaptic strength (Exc > Inh)
p.syn.J0_EI_sigma = 0.05; % std of J0 (Inh > Exc)
p.syn.J0_EE_sigma = 0.05; % std of J0 (Exc > Exc)
p.syn.J0_II_sigma = 0; % std of J0 (Inh > Inh)
p.syn.J0_IE_sigma = 0; % std of J0 (Exc > Inh)
p.syn.Jbar_EI = -2.4; % LTPi/LTDi switching point
p.syn.Jmax_EI = 0; % max I>E weights
p.syn.Jmax_EE = inf; % max E>E weights
p.syn.Jmax_II = 0; % max I>I weights
p.syn.Jmax_IE = inf; % max E>I weights
p.syn.Jmin_EI = -inf; % min I>E weights
p.syn.Jmin_EE = 0; % min E>E weights
p.syn.Jmin_II = -inf; % min I>I weights
p.syn.Jmin_IE = 0; % min E>I weights
p.syn.is_plastic = 1; % plasticity of synapses (all)
p.syn.is_plastic_EI = 1; % plasticity of synapses (Inh > Exc)
p.syn.is_plastic_EE = 1; % plasticity of synapses (Exc > Exc)
p.syn.is_plastic_II = 0; % plasticity of synapses (Inh > Inh)
p.syn.is_plastic_IE = 0; % plasticity of synapses (Exc > Inh)
% == theta dynamics ==
p.syn.g = 4.1; % coeffecient \gamma
p.syn.g_EI = 60; % coeffecient \gamma_{EI}
p.syn.g_EE = 114; % coeffecient \gamma_{EE}
p.syn.theta_L = -70; % resting value for \theta
p.syn.theta_rf = 0; % maximum value for \theta
p.syn.theta_fl = -140; % minimum value for \theta
p.syn.tau_theta = 12800; % decay time constant for \theta
% == exc learning rule ==
p.syn.b1 = 1.2e-5; % parameter in excitatory learning rule
% p.syn.b2 = 2.5; % parameter in excitatory learning rule
p.syn.b2 = -2.76e-4;% parameter in excitatory learning rule
p.syn.L2 = 2.1; % parameter in excitatory learning rule
p.syn.C2 = -1.4; % parameter in excitatory learning rule
p.syn.k2 = 1.5; % parameter in excitatory learning rule
p.syn.theta0_EE = -0.1; % parameter in excitatory learning rule
p.syn.V_ref_EE = -120; % parameter in excitatory learning rule
% == inh learning rule ==
p.syn.a1 = 0.00016; % parameter in inhibitory learning rule
p.syn.a2 = 0.0048; % parameter in inhibitory learning rule
p.syn.L1 = 1; % parameter in inhibitory learning rule
p.syn.C1 = -0.03; % parameter in inhibitory learning rule
p.syn.k1 = 1.5; % parameter in inhibitory learning rule
p.syn.theta0_EI = 0; % parameter in inhibitory learning rule
p.syn.V_ref_EI = -69; % parameter in inhibitory learning rule
% == external spikes/current ==
p.espk.type = 'constant'; % type of external input (constant/poisson)
p.espk.model_as = 'current'; % model as <current> (integrated) or <spike> (no integration)
p.espk.freq_E = 10; % frequency of external input (when method is set to poisson) for Exc neurons, in Hz
p.espk.freq_I = 10; % frequency of external input (when method is set to poisson) for Inh neurons, in Hz
p.espk.preEspk = 0; % time before delievery of external input, in ms
p.espk.dur = p.time.tmax-p.time.t0; % duration of delievery of external input, in ms
p.espk.gConn_E = 1.5; % connection strength to Exc neurons
p.espk.gConn_I = 1.5; % connection strength to Inh neurons
% == stimulus spikes/current ==
p.stim.typeStim = 'constant'; % type of stimulus: constant/poisson
p.stim.model_as = 'current'; % model as <current> (integrated) or <spike>
p.stim.preStim = 0; % time before the stimulus
p.stim.durStim = p.time.tmax-p.time.t0; % duration of stimulus
p.stim.freqStim = 20; % frequecy of expected poisson spike trains
p.stim.gConn = 1; % connection strength
p.stim.onoff = 1; % turn stimulus on/off
% == plot options ==
p.plot.plotDetails = 1; % plot detailed timeseries of each protocol (1: plot; 0: do not plot)
p.plot.plotEvery = 1; % plot every N round(s) of simulation
p.plot.plotPause = 5; % pause duration for plots to show (in sec)
p.plot.notify = 0; % notify when plots are ready (beep)
p.plot.closeUponShow = 0; % close plots after show
% == continued train ==
p.cont.placeholder = 0; % just a place holder for the field很长对吧,用它你能模拟1000个神经元的网络行为,但可能这种行为在真实人脑中完全不存在!
在太阳的核心,每秒钟将大约6亿吨氢聚变成氦,并在此过程中将400万吨物质转化为能量。这种能量可能需要10,000到170,000年才能逃离核心,是太阳的光和热的来源。当其核心的氢聚变减少到太阳不再处于流体静力平衡的点时,其核心将经历密度和温度的显著增加,同时其外层膨胀,最终将太阳转变为红巨星。据推算,太阳将变得足够大,可以吞噬水星和金星,并使地球无法居住。但这大约是五十亿年后的状况。在这之后,它将脱落其外层,裸露出核心,成为一种被称为白矮星的致密恒星。虽然不再通过核聚变产生能量,但仍会发光并散发出先前残余的热量。
上面的话的确是很有道理的。
自史前时代以来,太阳对地球的巨大影响就已得到认可。太阳被某些文化视为神。地球的自转及其围绕太阳的轨道是一些太阳历的基础。今天使用的主要日历是公历,它将基于16世纪观察到的太阳运动作为实际运动的标准来解释[23]。

我站在舞台中央
好似阳光洒在身上
数值模拟难题
数值模拟是啥哟,反正是挺复杂的一个东西,让我们分步来仔细的讲一讲吧!
太阳的数值模拟
为了更好的研究太阳对地球人类可能造成的影响,地球上的「高贵」、「自以为是」的人类们用所谓的一些人造的符号,极其丑陋和粗鄙地描述着太阳这一伟大造物的属性,就像这样:
或者是这样:
在可怜的人类眼里看来,把这些奇奇怪怪的符号添加到他们用沙子制造的某种「芯片」中,他们就能够一窥造物主的神力。有些不自量力的人类甚至认为他们因此便可以比肩造物主?可笑至极。在造物主看来,他们不过是产生热能罢了。
次级标题
在可怜的人类眼里看来,把这些奇奇怪怪的符号添加到他们用沙子制造的某种「芯片」中,他们就能够一窥造物主的神力。有些不自量力的人类甚至认为他们因此便可以比肩造物主?可笑至极。在造物主看来,他们不过是产生热能罢了。
再次级标题
在可怜的人类眼里看来,把这些奇奇怪怪的符号添加到他们用沙子制造的某种「芯片」中,他们就能够一窥造物主的神力。有些不自量力的人类甚至认为他们因此便可以比肩造物主?可笑至极。在造物主看来,他们不过是产生热能罢了。
再次级标题
在可怜的人类眼里看来,把这些奇奇怪怪的符号添加到他们用沙子制造的某种「芯片」中,他们就能够一窥造物主的神力。有些不自量力的人类甚至认为他们因此便可以比肩造物主?可笑至极。在造物主看来,他们不过是产生热能罢了。
最次级的标题
在可怜的人类眼里看来,把这些奇奇怪怪的符号添加到他们用沙子制造的某种「芯片」中,他们就能够一窥造物主的神力。有些不自量力的人类甚至认为他们因此便可以比肩造物主?可笑至极。在造物主看来,他们不过是产生热能罢了。
这是一个极度简化,或者说极度简陋的模型
这个模型再简单不过,就是一个强制把抑制性突触更新规则与兴奋性突触更新规则进行偶联的模型,放在这除了博人一笑,就仅仅是为了测试详细信息块的样式了。
| 参数 | 数值 | 描述 |
|---|---|---|
| A | 10 | 公式1中系数A的值 |
| B | 10.3 | 公式1中系数B的值 |
| C | 1.8 | 公式1中系数C的值 |
| 页脚1 | 页脚2 | 页脚3 |
写了这么多无意义的表格,我不禁诗兴大发,来吧,作诗一首:
当你老了,头白了,睡意昏沉,
炉火旁打盹,请取下这部诗歌,
慢慢读,回想你过去眼神的柔和,
回想它们昔日浓重的阴影;
多少人爱你青春欢畅的时辰,
爱慕你的美丽,假意或真心,
只有一个人爱你那朝圣者的灵魂,
爱你衰老了的脸上痛苦的皱纹;
垂下头来,在红光闪耀的炉子旁,
凄然地轻轻诉说那爱情的消逝,
在头顶的山上它缓缓踱着步子,
在一群星星中间隐藏着脸庞。
多媒体
这个板块我主要想试试所有的多媒体组件,看看他们的表现是否真的符合我的预期,希望一切顺利吧,祝我好运🍀!
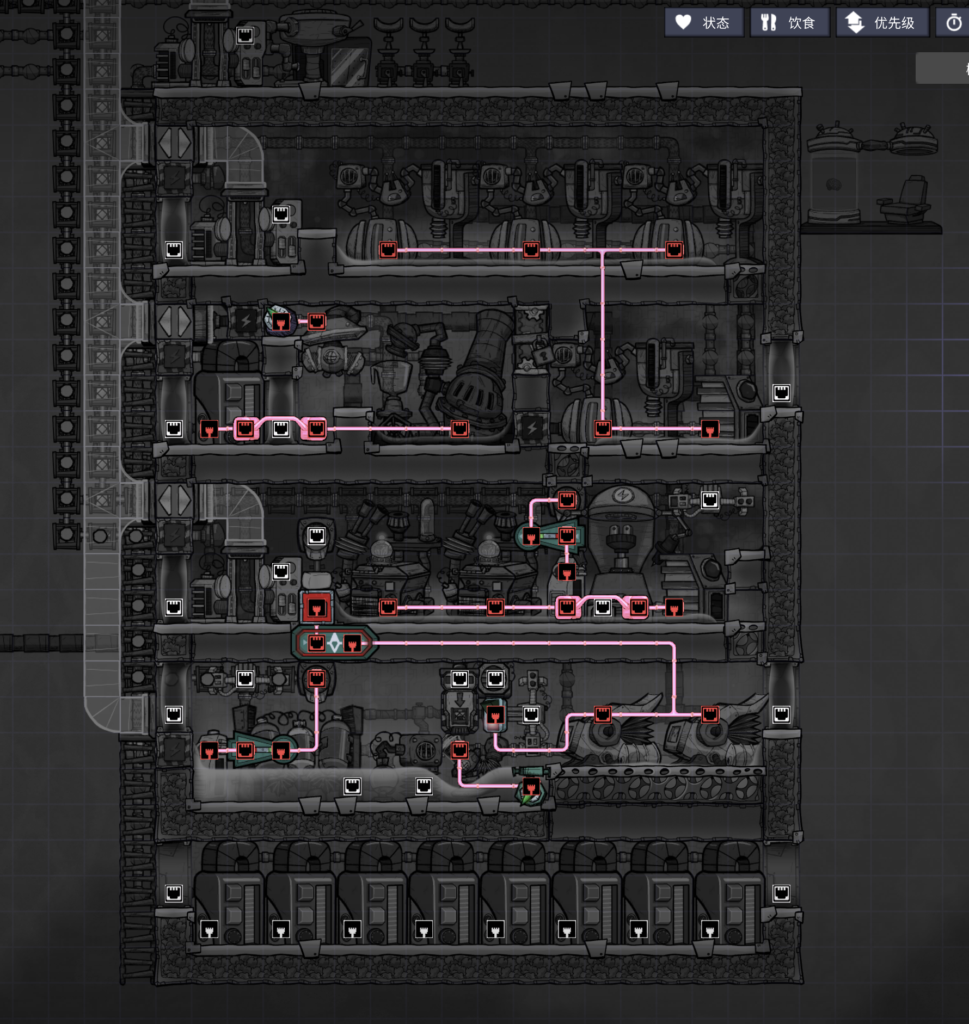
这个是我「缺氧」鸟托邦里建设基地的化石能源发电厂的信号线连接图,那个悬空的震荡仪不重要不用理会。
再来听一听音乐吧:
骗你的上面这个其实是一个人在念一个名为「天鹅」的词语,还是来点正经的音乐吧:
嵌入元素的测试
总算是搞完了,现在测试一下列表的样式:
第一步:初始化项目
- 运行
npm init -y - 配置
.gitignore文件 - 安装基础依赖:
- TypeScript
- ESLint
- Prettier
- 哈哈哈哈啊哈
包含内联代码的列表:
- 使用
flex-direction: column;来实现垂直布局。 - 点击
<a>链接跳转到 测试页面。
发现一篇挺有意思的文章,由彼得所著,风景美不胜收:
按键元素
我想测试一下按键组,请选择点击以下按键:
这是横向排列的按钮:
这两个是纵向排列的
嗯,没错,上面是一些测试用的按键。
“何谓太阳” 的回复
什莫啊?
你就是个粪蛋子,不是什么太阳国王哈!
粪蛋子是我,我就是粪蛋子💩
太阳穴是热点哈
盲生,你发现了华点!
国王喊我发两条评论,是王下发的号令!执行!
多一条不行,少一条不可!